





Введение
Домашняя кошка (Felis catus L.) является одной из интереснейших моделей для изучения микроэволюционных процессов. Это связано с тем, что у нее имеется ряд генов, ответственных за окрас и структуру меха, а эти мутантные гены, в свою очередь, достаточно просто анализировать визуально. Все цветовые вариации в окрасе меха у кошек F. catus, кроме серого полосатого (дикий тип), обусловлены мутациями в генах, контролирующих процесс пигментогенеза и миграции меланоцитов в волосяные фолликулы. Частоты встречаемости мутантных и нормальных аллелей генов окраса характерны для каждой популяции и отражают их генетические структуры. В настоящее время аллельные частоты по генам окраса меха домашних кошек описаны в основном для всех развитых стран. На территории Беларуси такие исследования практически не проводились, поэтому работа направленная на изучение генетической структуры F. catus городов Беларуси крайне актуальна [1–2].
Цель работы: определить генетическую структуру популяций F. catus востока Беларуси, а также используя полученные данные провести оценку их генетической дифференциации.
Материалы иметоды
Для достижения поставленной цели были исследованы 7 городов (Осиповичи, Бобруйск, Чаусы, Костюковичи, Славгород, Шклов, Могилев) восточной части Беларуси. В каждом городе был проложен маршрут, на котором фиксировались все встреченные особи F. catus.
Сбор материала проводился с 8:00 до 10:00 утра и с 20:00 до 22:00 вечера, так как именно в это время суток у особей F. catus наблюдается повышенная активность [3].
Для каждой встреченной особи был присвоен свой номер и составлен ее генетический портрет. Всего таким образом было проанализировано 982 особи F. catus.
Установление генотипов осуществлялось методом визуального типирования характера и окраски шерстного покрова животных.
Определение частот встречаемости мутантных аллелей проводилось по 8 генам окраса и структуры шерсти: сцепленного с полом гена Orange (доминантный аллель О), а также семи аутосомных генов (Agouti — рецессивный аллель а, Dilute — рецессивный аллель d, Long hair — рецессивный аллель l, Piebald spotting — доминантный аллель S, White — доминантный аллель W, Tabby — рецессивный аллель tb, Color — рецессивный аллель cs). Все мутантные аллели, за исключением аллеля l, влияют на окраску шерстного покрова и характер его распределения. Аллель l в гомозиготе определяет длинную шерсть. Фенотипическое проявление аллелей, их взаимодействие и полный расчет аллельных частот подробно описаны ранее [2, 4–6].
Визуализация распределения аллельных частот в исследованных городах на географической карте проводилась с помощью геоинформационного пакета Golden Software Surfer v.9.9.
Результаты иих обсуждение
Для каждого из 7 исследованных городов востока Беларуси была определена генетическая структура популяций F. catus по 8 генам окраса и структуры шерсти. Полученные результаты были сведены в таблицу 1.
Таблица 1
Генетическая структура популяций F. catus исследованных городов
|
Популяция |
Частота мутантного аллеля |
|||||||
|
a |
tb |
d |
l |
S |
W |
cs |
O |
|
|
Осиповичи |
0,724 ±0,038 |
0 |
0,484 ±0,045 |
0,598 ±0,041 |
0,228 ±0,033 |
0,005 ±0,005 |
0,103 ±0,051 |
0,202 ±0,037 |
|
Бобруйск |
0,806 ±0,030 |
0,137 ±0,068 |
0,478 ±0,041 |
0,540 ±0,038 |
0,250 ±0,031 |
0,025 ±0,010 |
0,132 ±0,046 |
0,223 ±0,035 |
|
Чаусы |
0,729±0,027 |
0,104±0,052 |
0,461±0,034 |
0,530±0,031 |
0,234±0,024 |
0,019±0,007 |
0,200±0,037 |
0,227±0,028 |
|
Костюковичи |
0,728±0,034 |
0 |
0,429±0,042 |
0,454±0,040 |
0,205±0,028 |
0,029±0,011 |
0,296±0,045 |
0,175±0,031 |
|
Славгород |
0,774±0,028 |
0,176±0,061 |
0,368±0,039 |
0,475±0,037 |
0,254±0,027 |
0,007±0,005 |
0,224±0,041 |
0,154±0,027 |
|
Шклов |
0,766±0,036 |
0,194±0,067 |
0,374±0,045 |
0,569±0,039 |
0,233±0,031 |
0,018±0,009 |
0,167±0,048 |
0,192±0,034 |
|
Могилёв |
0,705±0,027 |
0,134±0,047 |
0,399±0,032 |
0,470±0,030 |
0,331±0,026 |
0,026±0,008 |
0,141±0,035 |
0,179±0,024 |
Из таблицы видно, что наиболее высокие частоты были встречены для следующих аллелей а, d и l, значения которых во всех исследованных популяциях варьировали от 37 до 81 %. Мутантные аллели S, O и cs характеризовались средним значением частот встречаемости, в большинстве исследованных популяций имели величины от 10 до 33 %. Мутантные аллели W, tb встречались с низкой частотой или отсутсвовали.
Высокое значение частоты мутантного аллеля а вызвано плейотропным влиянием гена окраса на поведение. Этот мутантный аллель а можно назвать аллелем урбанизации. Его частота возрастает прямо пропорционально росту численности населения. Поэтому черные кошки более резистентны к стрессу городской среды, чем их собратья с окрасом «дикого типа» [2, 7].
Что касается низкой частоты встречаемости аллеля tb, а в некоторых популяциях его полного отсутствия это связано с тем, что данный аллель характерен для популяций Западной Европы, где он исторически возник и частота его достигает 80–90 %.
На втором этапе исследований была проведена оценка генетической дифференциации популяций F. catus исследованных городов.
Существенные отличия были встречены только для двух аллелей: рецессивного аллеля l и доминантного аллеля S.
Частота аллеля l заметно увеличивается в направлении с юга на север (рисунок 1), что подтверждается данными полученными ранее [8, 9]. По этим данным в более северных районах кошки с длинной шерстью встречаются чаще, чем с короткой.
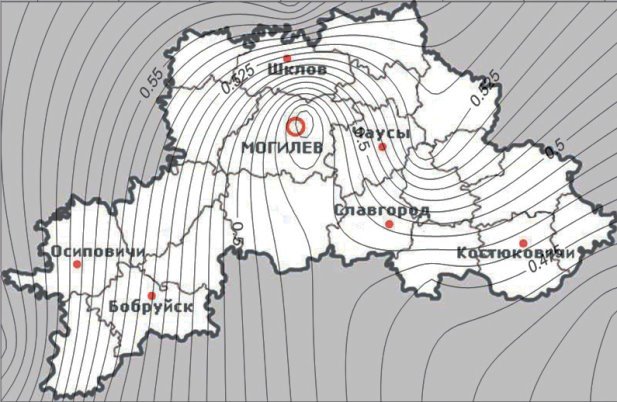
Рис. 1. Карта распространения кошек с мутацией l
Ситуация с частотой аллеля S оказалась несколько иной. С максимальным значением аллель пегости был встречен в Могилеве. По мере удаления от областного центра к периферии частота данного аллеля в исследованных городах оказывалась ниже (рисунок 2), чего нельзя сказать о городах более удаленных от Могилева (Гомель, Орша), но имеющих схожие аллельные частоты [8]. Это можно связать с проходящими через эти города железнодорожными сообщениями, что способствует более частой миграции людей в направлении Орша-Могилев-Гомель и обратно, чем от Могилева к периферийным городам.
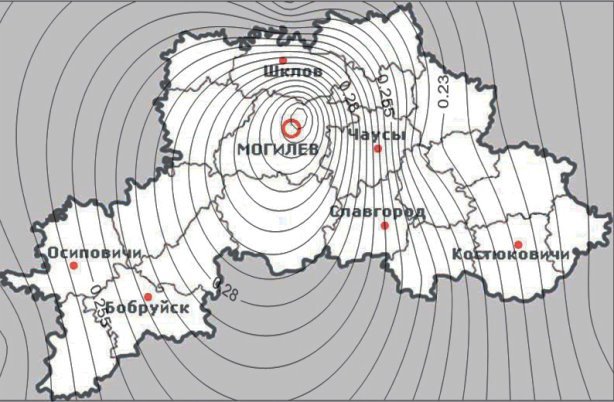
Рис. 2. Карта распространения кошек с мутацией S
Заключение
В ходе проведенных исследований были получены следующие результаты:
- составлены генетические портреты особей F. catus, обитающих в 7 городах (Осиповичи, Бобруйск, Чаусы, Костюковичи, Славгород, Шклов, Могилев) восточной части Беларуси;
- определена генетическая структура популяций F. catus по 8 генам окраса и структуры шерсти;
- проведена оценка генетической дифференциации популяций F. catus обитающих в 7 городах восточной части Беларуси.
Работа проводилась в рамках тем ГПНИ 16–14 и ГПНИ 16–32, выполняемых в рамках Государственных программ «Биотехнологии» и «Природопользование и экология».
Авторы выражают признательность чл.-корр. НАНБ, д.б.н., профессору Г. Г. Гончаренко за помощь и поддержку в популяционно-генетических исследованиях, а также консультации во время написания данной работы.
Литература:
1 Christensen, A. C. Cats as an Aid to Teaching Genetics / A. C. Christensen // Genetics. — 2000. — 155. — P. 999–1004.
2 Гончаренко, Г. Г. Генетика. Анализ наследственных закономерностей на генах меха кошек Felis catus / Г. Г. Гончаренко, С. А. Зятьков. — Гомель: ГГУ им. Ф. Скорины, 2007. — 108 с.
3 Антоненко, Т. В. Методика оценки территориальной активности Felis catus в антропогенных условиях обитания / Т. В. Антоненко // Вестник Алтайского государственного аграрного университета, 2009 — № 10 (60). — С. 110–113.
4 Robinson, R. Genetics for Cat Breeders / R. Robinson. — Oxford: Pergamon, 1991. — 234 p.
5 Гончаренко, Г. Г. Мутантные гены окраски в популяциях домашних кошек Средней Азии и Европейской части СССР / Г. Г. Гончаренко, О. Е. Лопатин, Г. П. Манченко // Генетика, 1985. — Т. ХХI. — № 7. — С.1151–1158.
6 О'Брайен, С. Генетика кошки / С. О'Брайен, Р. Робинсон, А. С. Графодатский, А. В. Таранин. — Новосибирск: Наука, 1993. — 212 с.
7 Бородин, П. М. Кошки и гены / П. М. Бородин. — М.: Книжный дом ЛИБРОКОМ, 2011. — 136 с.
8 Гончаренко, Г. Г. Уровень генетической дифференциации у кошек Felis catus (L.) в западноевропейских, североамериканских и восточноевропейских популяциях / Г. Г. Гончаренко, С. А. Зятьков // Вавиловский журнал генетики и селекции. — Новосибирск: ИЦиГ СО РАН. — Т. 15, № 3, 2011. — С. 516–523.
9 Ruiz-Garcia, M. Is There Really Natural Selection Affecting the l Frequencies (Long Hair) in the Brazilian Cat Populations? / M. Ruiz-Garcia // The Journal of Heredity, 2000. V. –91(1). P. — 49–57.
