





С целью выяснения видовых особенностей строения структур носовой области человека и некоторых млекопитающих в пренатальном периоде онтогенеза, исследованы 75 препаратов передплодов и плодов человека, 85 препаратов млекопитающих разных периодов пренатального развития с использованием комплекса методов морфологического исследования (антропометрия, морфометрия, микроскопия, трехмерное компьютерная реконструкция, статистический анализ). Установлено, что видовые различия строения носового аппарата изученных млекопитающих связаны с формой носовой полости, структурой вторичного костного неба, количеством и степенью развития носовых раковин, околоносовых пазух и обусловлены особенностями питания, характером зубочелюстной системы, степенью развития головного мозга, интенсивностью дыхания и условиями существования животного. У человека латеральная носовая железа отсутствует, но хорошо развиты респираторные, обонятельные и верхнечелюстные железы.
Ключевые слова: носовая область, сравнительный эмбриогенез, млекопитающие, человек
Сравнительно-анатомические и сравнительно-эмбриологические исследования не теряют своей актуальности в изучении строения органов и систем организма человека для выяснения закономерностей морфогенеза органов и систем организма различных животных, определение общих черт и особенностей морфо-функциональной организации живой материи [1, с. 289–292; 2, с. 454–461; 3, с. 77–81; 4, с. 314–317]. Среди позвоночных животных именно у млекопитающих носовая полость достигла наивысшего своего развития, и кроме воздухоподводящей функции, иммунной, защитной, участвует в процессах терморегуляции и хеморецепции [5, с. 273–284; 6, с. 281–293; 7, с. 1325–1340]. Анализ современной научной литературы позволяет прийти к выводу, что проблема пренатального морфогенеза структур носовой полости в сравнительно-эмбриологическом плане остается нерешенной [8, с. 2065–2079; 9, с. 37–62]. В связи с этим существует необходимость комплексного методологического подхода к изучению строения и развития носового аппарата в сравнительном плане у разных видов млекопитающих в связи с местом и способом их существования [1, с. 289–292; 10, с. 137–141].
Цель исследования — определить видовые особенности строения структур носовой области человека и некоторых млекопитающих в пренатальном периоде онтогенеза.
Материалы иметоды. Исследование проведено на 75 препаратах зародышей и передплодов человека и 85 препаратах млекопитающих разных периодов пренатального развития. Изучено 5 видов, относящихся к 4 рядам класса млекопитающих: человек, белая крыса, кошка домашняя, собака домашняя, свинья домашняя. Животные для сравнительно-анатомического исследования подбирались с учетом условий их существования, особенностей питания, размножения, степени развития головного мозга и органов чувств. Материал (зародыши и передплоды человека) получали из Черновицких акушерско-гинекологических клиник. Серии гистологических срезов зародышей млекопитающих, зародышей и передплодов человека взяты из музея кафедры анатомии человека им. М. Туркевича и учебно-научного фонда кафедры гистологии, цитологии и эмбриологии Высшего государственного учебного заведения Украины «Буковинский государственный медицинский университет».
Мы воспользовались разделением пренатального периода онтогенеза млекопитающих на 16 последовательных стадий (Тятенкова Н. Н., 1998) [10, с. 137–141]. Несмотря на то, что видовые особенности морфогенеза отчетливо проявляются на протяжении большого органогенеза, описание этого периода мы начали с третьей стадии.
Таблица 1
Соответствие стадий развития млекопитающих, ТКД (мм)*
|
Номер стадии |
человек |
кошка домашняя |
крыса белая |
собака домашняя |
Свинья домашняя |
|
3 |
4,0–6,0 |
4,0 |
10 |
4,0–5,0 |
- |
|
4 |
6,0 |
5,0 |
11 |
6,0–8,0 |
- |
|
5 |
7,0–11,0 |
6,0 |
12 |
9,0 |
- |
|
6 |
12,0–13,0 |
8,5–9,0 |
13 |
10,0 |
9,0–13,0 |
|
7 |
14,0–15,0 |
10,0–11,0 |
14 |
11,0–12,0 |
14,0–15,0 |
|
8 |
16,0–17,0 |
11,0–12,0 |
15 |
13,0–15,0 |
16,0–18,0 |
|
9 |
18,0–20,0 |
14,0–17,0 |
15 |
16,0 |
19,0–23,0 |
|
10 |
21,0–24,0 |
18,0–21,0 |
16 |
17,0–19,0 |
24,0–27,0 |
|
11 |
25,0–29,0 |
22,0–23,0 |
16 |
22,0–28,0 |
28,0 |
|
12 |
30,0–38,0 |
25,0–27,0 |
16–17 |
28,0–39,0 |
33,0–34,0 |
|
13 |
39,0–79,0 |
28,0–35,0 |
17 |
30,0 |
36,0–50,0 |
|
14 |
80,0–189,0 |
36,0–59,0 |
18–19 |
31,0–69,0 |
51,0–54,0 |
|
15 |
190,0–279,0 |
60,0–100,0 |
20–21 |
70,0 |
55,0–150,0 |
|
16 |
280,0–370,0 |
101,0–120,0 |
22 |
140,0 |
- |
Примечание: * для крысы белой возраст приведен в сутках.
Исследование проведено на препаратах зародышей, передплодов и плодов человека и млекопитающих без патологии носовой области.
Результаты исследования иих обсуждение. В конце плодного периода онтогенеза носовая область у млекопитающих изученных видов приобретает черты дефинитивного строения. Она окружена хрящевой носовой капсулой, к внешней поверхности которой прилегают: дорсально — носовые и, частично, лобные, латерально — верхнечелюстные кости. От полости черепа носовая полость отделена решетчатой пластинкой клиновидной кости, которая состоит из хрящевой ткани. Носовая полость разделена на две половины хрящевой носовой перегородкой, которая аборально переходит в хрящевую основу черепа, дорсально — в свод носовой капсулы (рис. 1). Вентральную стенку носовой полости образуют парасептальные хрящи и вторичное небо.
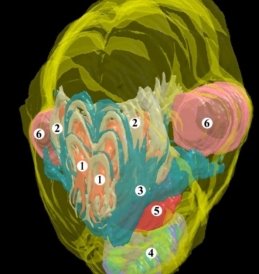
Рис. 1. Трехмерная компьютерная реконструкция головы крысы 21,0 мм ТКД. Передне-левая проекция. Ув. 1:12; 1 — носовая полость; 2 — хрящевая капсула; 3 — костная ткань, верхняя челюсть; 4 — нижняя челюсть; 5 — язык; 6 — глазные яблоки
У новорожденных белой крысы носовая полость удлиненная дорсо-вентрально и занимает 45 % длины лицевого черепа (рис. 2), в оральном отделе она овальной формы на поперечном срезе, а в средней части — треугольной. Хрящевая носовая капсула хорошо развита. Максиллярная раковина простой формы, валикообразная.
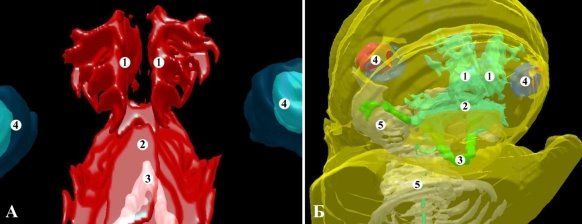
Рис. 2. Трехмерная компьютерная реконструкция головы крысы 19,0 мм ТКД (А) и 21,0 мм ТКД (Б). А — верхне-задняя проекция, Б — передне-правая проекция. Ув. 1:12; 1 — носовая полость; 2 — ротовая полость; 3 — меккелев хрящ; 4 — глазные яблоки; 5 — зачаток скелета и костей черепа
Назотурбиналия проходит вдоль дорсо-латеральной стенки и имеет хрящевую опорную часть (рис. 3). Этмоидальные раковины делятся на внешние и внутренние. Общий носовой ход в переднем отделе сужен, в среднем — резко расширяется. Хорошо развит решетчатый лабиринт и пазухи решетчатой кости. Нижний носовой ход не выражен, средний носовой ход достаточно широк.
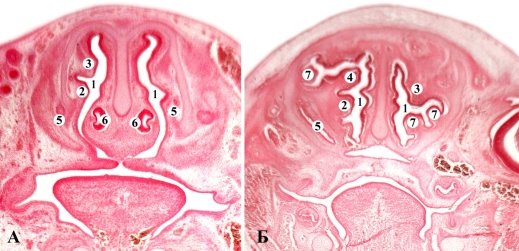
Рис. 3. Фронтальные срезы носовой участка крысы 18,0 мм ТКД. Гематоксилин-эозин. Микрофото. Об. 8, ок. 7: 1 — полость носа, 2 — максиллотурбиналия; 3 — назотурбиналия; 4 — эндо-турбиналии; 5 — носослезный канал; 6 — Лемешев-носовой орган; 7 — экзо-турбиналии
У белой крысы в конце внутриутробного развития хорошо развиты железы носовой полости. Обнаружены закладки верхнечелюстных, латеральной, передних медиальных, респираторных и обонятельных желез. Латеральная носовая железа крысы хорошо развита, по строению является сложной железой альвеолярного типа с разветвлённым конечным отделом. От основной протоки железы отходят протоки второго и третьего порядков, слизистая оболочка в области конечного отдела железы сильно утолщена. Передних медиальных желез к рождению две. Располагаются они параллельно друг другу в слизистой оболочке носовой перегородки. Аборальный отдел желез спиралеобразно закрученный.
У свиньи домашней пятнадцатой стадии внутриутробного развития носовая полость удлиненная и занимает 52 % длины черепа, ее передний отдел сужен и содержит Т-образную максиллярную раковину (рис. 4).
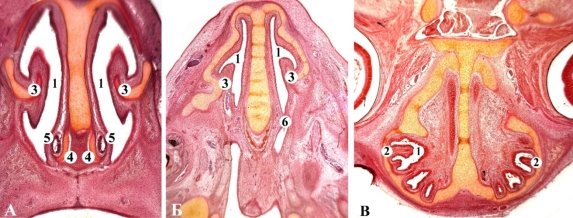
Рис. 4. Горизонтальные (А, Б) и фронтальной (В) срезы носовой полости зародышей поросенка 44,0 мм ТКД. Гематоксилин-эозин. Микрофото. Об. 8, ок. 7: 1 — носовая полость; 2 — этмоидальные раковины 3 — максиллярная раковина; 4 — парасептальный хрящ; 5 — лемешево-носовой орган; 6 — носонебный канал
Дорсо-латерально находится небольшая, с нечетко выраженной опорной частью, валикообразная назотурбиналия. Носовая перегородка массивная, равномерно утолщенная по всей длине. Вторичные хоаны расположены в задней трети носовой полости и ведут в носоглоточный ход. Передняя поперечная пластинка орально срастается с нижним краем носовой перегородки, аборально остается свободной, размещаясь почти горизонтально. Надперегородочная борозда глубокая. Хорошо развитые парасептальные хрящи. Передний парасептальный хрящ в виде вертикальной пластинки, орально соединён с передней поперечной пластинкой, аборально — с носонебным хрящом. Последний выстилает одноименный канал. В аборальном направлении от носонебного хряща отходит короткий задний парасептальный хрящ в виде горизонтальной пластинки. С носонебным хрящом соединен лемешево-носовой хрящ кольцеобразной формы и соединяется дорсально с передним парасептальным хрящом. Аборальный отдел носовой полости расширен и включает пять эндо-турбиналий разветвленной формы. В состав вторичного костного неба входит нижнечелюстная, верхнечелюстная и небная кости. Лемеш имеет вид нечетной закладки, прилегающей к вентральному краю носовой перегородки. Из околоносовых пазух развивается верхнечелюстная и лобная. Окостенения обнаружено в закладках неба и лемеша.
У кошки домашней в конце плодного периода онтогенеза носовая полость укорочена и занимает 38 % длины черепа. Общий носовой ход в переднем отделе невысок. Средний отдел носовой полости шире, увеличивается к верху и содержит максилотурбиналию. Раковина хорошо развита, имеет простую, незначительно закрученную вниз форму (рис. 5 А, Б). От дорсо-латеральной стенки отходит небольшая валикообразная назотурбиналия. Аборальный отдел содержит ряд обонятельных раковин: 4 эндо- и 5 экто-турбиналий. Свободные края внутренних раковин раздваиваются и образуют завитки (рис. 5 В). Большинство этмоидальных раковин отходят от латеральной стенки носовой капсулы, часть — от дорсальной.
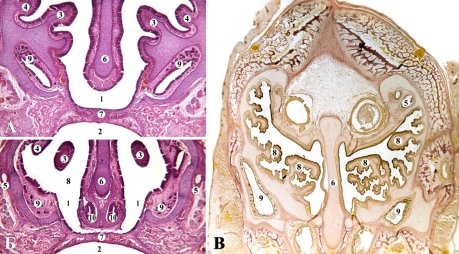
Рис. 5. Фронтальные срезы новорожденных кошки. А, Б — гематоксилин-эозин, В — Ван Гизону. Микрофото. Об. 8, ок. 7: 1 — носовая полость; 2 — ротовая полость; 3 — максиллярная раковина; 4 — назотурбиналия; 5 — носослезный канал; 6 — носовая перегородка; 7 — вторичное костное небо; 8 — этмоидальные раковины 9 — латеральная носовая железа; 10 — лемешево-носовой орган
Верхнечелюстная пазуха хорошо развита, ее слепой отдел заканчивается в аборальном отделе носовой полости. На поперечном срезе пазуха вытянута дорсо-вентрально. Вторичное костное небо формируют парные закладки межчелюстной, верхнечелюстной, небной костей и лемеш. Последний входит в состав костного неба в виде небольшого вкрапления. Лемеш представлен в оральном отделе в виде двух вертикальных пластин, расположенных между органом Якобсона. В аборальном пластинки объединяются, и нечеткая закладка охватывает вентральный край хрящевой носовой перегородки. Нижняя половина носовой перегородки значительно расширена в области лемешево-носового органа. Надперегородочная борозда хорошо выражена. Передняя поперечная пластинка занимает горизонтальное положение, но в аборальном направлении она приобретает подковообразною форму и срастается с носовой перегородкой. Передний парасептальный хрящ в виде самостоятельной вертикальной пластинки. Носонебный хрящ сопровождает одноименный канал и имеет форму изогнутой вертикальной пластинки, соединенной с лемешево-носовым хрящом. Последний в области выводящей протоки органа Якобсона имеет кольцеобразную форму, в аборальном — желобовидную. Окостенения обнаружено в опорных частях максиллярных и этмоидальных раковин, в аборальном отделе носовой перегородки и закладка вторичного костного неба.
У новорожденных кошки домашней латеральная носовая железа по строению является простой альвеолярной железой с относительно короткой выводящей протокой и разветвленным конечным отделом, развита умеренно. Слизистая оболочка в области конечного отдела железы умеренно утолщена. Передние медиальные железы отсутствуют. В переднем отделе носовой полости слизистая оболочка содержит большое количество респираторных желез, которые по строению являются простыми альвеолярными железами с разветвленным конечным отделом. В верхнечелюстной пазухе количество желез резко увеличивается, они приобретают более сложную, трубчастую альвеолярную форму. В околоносовых пазухах обнаружена верхнечелюстная и лобная пазухи.
У новорожденных собаки домашней носовая полость удлиненная. В крыше носовой капсулы отмечена глубокая надперегородочная борозда. Нижний край носовой перегородки немного утолщенный. Максиллярная раковина в виде сложной пространственной формы, ее свободный край характеризуется складчатым строением, что значительно увеличивает площадь внешней поверхности (рис. 6 А). Назотурбиналия небольшая, с хрящевой опорной частью. Обонятельные раковины делятся на внутренние и внешние, приобретая сложную разветвленную и закрученную формы. Эндотурбиналий насчитывается четыре, экзотурбиналий — шесть. В парасептальных хрящах обнаружены передний парасептальный, носонебный, хрящ небного сосочка и С-образный лемешево-носовой. Лемеш представлен в виде нечетной закладки V-образной формы. Вторичное костное небо образовано закладками верхнечелюстной, межчелюстной и небной костей. Из дополнительных пазух присутствует верхнечелюстная и лобная. Центры окостенения обнаружены в закладках лемеша, верхнечелюстной, межчелюстной, небной костей, максиллярной и этмоидальной раковинах.
У новорожденных собаки домашней хорошо развиты железы носовой полости (рис. 6 Б): респираторные, верхнечелюстные, латеральная железы. Латеральная носовая железа собаки — это простая альвеолярная железа с разветвленным конечным отделом, слизистая оболочка в области конечного отдела немного утолщена. С околоносовых пазух развиты верхнечелюстные и лобные.
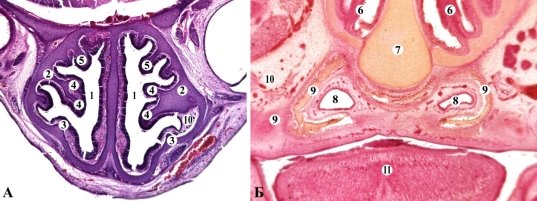
Рис. 6. Фронтальные срезы зародыша собаки 12,0 мм ТКД. Гематоксилин-эозин. Микрофото. Об. 8, ок. 7: 1 — носовая полость; 2 — верхнечелюстные раковины 3 — носовые раковины 4 — эндотурбиналии; 5 — экзотурбиналии; 6 — этмоидотурбиналии; 7 — лемеш; 8 — лемешево-носовой орган; 9 — лемешево-носовой хрящ; 10 — язык
У плодов человека тринадцатой стадии развития носовая капсула, раковины и носовая перегородка состоит из хрящевой ткани. От латеральных стенок носовой полости отходят носовые раковины: нижняя, средняя и верхняя. Их свободные, медиальные края направлены вентро-медиально. Все раковины имеют простую без завитков форму (рис. 7). В переднем отделе носовая полость щелевидной формы, удлиненная в дорсо-вентральном направлении. В среднем отделе она расширена, на поперечном разрезе ромбовидной формы. Нижний носовой ход сохраняется преимущественно в виде эпителиального тяжа, средний ход короткий, широкий; верхний имеет вид углубления, которое простирается в дорсо-латеральном направлении. В околоносовых пазухах присутствует небольшая клиновидная и верхнечелюстная. Вторичное небо в форме свода, в его состав входят парные закладки верхне-челюстной и небной костей. В аборальном над последними расположена нечетная подковообразная закладка лемеша, в котором определяется три центра окостенения. У человека латеральная носовая железа отсутствует, хорошо развиты респираторные, обонятельные и верхнечелюстные железы.
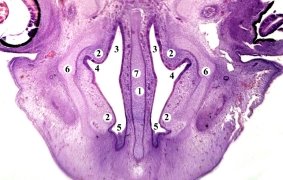
Рис. 7. Фронтальный срез носовой участка передплодов 25,0 мм ТКД. Гематоксилин-эозин. Микропрепарат. Об. 8, ок. 7: 1 — носовая перегородка; 2 — носовые раковины 3 — верхний носовой ход; 4 — средний носовой ход; 5 — нижний носовой ход; 6 — хрящевая капсула
Выводы:
1. Видовые различия в строении носового аппарата изученных млекопитающих связаны по форме носовой полости, строением вторичного костного неба, количеством и степенью развития носовых раковин, околоносовых пазух, и обусловлены особенностями питания, характером зубочелюстной системы, степенью развития головного мозга, интенсивностью дыхания и условиями существования животного.
2. У человека латеральная носовая железа отсутствует, хорошо развиты респираторные, обонятельные и верхнечелюстные железы.
Перспективы дальнейших исследований. Перспективным направлением сравнительно-эмбриологических исследований — установление особенностей источников кровоснабжения и иннервации носовой области в пренатальном периоде онтогенеза человека и некоторых животных.
Литература:
- Кузняк Н. Б. Функциональное значение некоторых анатомических образований внутреннего носа и их развитие у животных в фило- и онтогенезе / Н. Б. Кузняк, С. М. Шувалов // Молодой учëный. — 2016. — № 6.
- Kuzniak N. B. Development of the inner nasal cavity in animals in phylo- and ontogenesis: functional anatomic significance in the development period / N. B. Kuzniak // Oxford Review of Education and Science. — 2016. — № 1.
- Шаповалова Е. Ю. Возрастная динамика формирования челюстно-лицевого аппарата человека в раннем периоде пренатального развития / Е. Ю. Шаповалова, А. Н. Барсуков, Г. А. Юнси // Морфология. — 2010. — Т. 137, № 2.
- Слободян О. М. Закономірності перинатальних органометричних параметрів ділянок і структур голови / О. М. Слободян, Н. Б. Кузняк, Л. П. Лаврів // Вісник проблем біології та медицини. — 2016. — № 2.
- Numerical modeling of odorant uptake in the rat nasal cavity / G. C.Yang, P. W. Scherer, K. Zhao [et al.] // Chemical senses. — 2007. — № 32.
- Respiratory turbinates of canids and felids: a quantitative comparison / B. Van Valkenburgh, J. Theodor, A. Friscia [et al.] // Journal of Zoology. — 2004. — № 264.
- Reconstruction and morphometric analysis of the nasal airway of the dog (Canis familiaris) and implications regarding olfactory airflow / B. A. Craven, T. Neuberger, E. G. Paterson [et al.] // The Anatomical Record. — 2007. — № 290.
- Respiratory and olfactory turbinals in feliform and caniform carnivorans: the influence of snout length / B. Van Valkenburgh, B. Pang, D. Bird [et al.]. // The Anatomical Record. — 2014. — № 297.
- Smith T. Anatomy of the nasal passages in mammals / T. Smith, T. Eiting, K. Bhatnagar // Handbook of Olfaction and Gustation. — 2015.
- Тятенкова Н. Н. Периодизация пренатального онтогенеза млекопитающих // Н. Н. Тятенкова. — Российские морфологические ведомости. — 2000. — №. 1–2.
