





Рассматриваются сроки появления и пребывания в планктоне личинок тихоокеанской мидии, динамика численности и размерного состава течение всего периода нахождения в планктоне в различных районах Тауйской губы. В связи с интенсивным прогревом вод в июле, массовый выход личинок мидий в планктон отмечен в конце июля - начале августа 2009 г. Нерест был растянут, личинки мидии встречались в планктоне с 25 июня по 20 сентября.
Ключевые слова: двустворчатый моллюск, личинки, велигеры мидии, численность, температура воды, места обитания.
Time of emergence and terms of stay in plankton of the Pacific mussel veligers was studied. Their population dynamics and compositional size during all period of staying in plankton in different areas of Tauisk Bay were analyzed. Due to intense warming of waters in July mass emergence of veligers into plankton occurred at the end of July – the beginning of August, 2009. Spawning had protracted terms, mussel veligers occurred in plankton since 25 of June till 20 of September.
В связи с возрастающим значением морских беспозвоночных в качестве объектов питания человека и сокращением их естественных запасов возрастает актуальность проблемы поиска способов их восстановления и пополнения. Во всем мире эта проблема успешно решается путем искусственного разведения и выращивания гидробионтов в море с применением различных технологий [14].
При создании хозяйств марикультуры в том или ином районе Тауйской губы возникает необходимость в биологическом и экологическом обосновании целесообразности выбора района для размещения плантации мидий. Важным этапом такого обоснования особенно для культивирования в подвесных коллекторах, является оценка состояния естественного воспроизводства мидий, о чем можно судить по результатам планктонных исследований. Известно, что формирование новых поселений осуществляется путём непосредственного осаждения личинок из планктона, а пополнение стационарных поселений – через вторичное оседание ювенальных особей [4].
Пелагические личинки двустворчатых моллюсков в Тауйской губе – является одним из часто встречающихся компонентом летнего меропланктона. Основным фактором, определяющим сроки присутствия личинок донных беспозвоночных в пелагиали, является корреляция сезонов размножения их донных родительских форм с температурой воды и местными сезонными циклами развития фитопланктона, поскольку основную долю состава пищи личинок донных беспозвоночных составляет фитопланктон [12]. Этот фактор определяет место обитания основной массы личинок – поверхностный десятиметровый слой водной толщи, а частый механизм запуска нереста – прогрев воды до определенной температуры в зависимости от места обитания родительских форм – в воде, на поверхности грунта, на литорали или верхней сублиторали [2]. Кроме того, установлено, что пополнение молодью литоральных популяций мидий крайне нестабильно и зависит от абиотических факторов среды. Одним из таких факторов, влияющих на выживание молоди на литорали является суммарная продолжительность осушения в период осаждения личинок [1].
Целью исследований было изучение численности, размерного состава и сроков пребывания в планктоне пелагических личинок тихоокеанской мидии в прибрежной части Тауйской губы в районах: м. Нюкля, бух. Нагаева и Старая Веселая в течение вегетационного сезона 2009 г. и определение наиболее перспективных районов для культивирования мидий.
Материал и методика
Исследования меропланктона проводились в период с 15 июня по 5 октября 2009 г. в прибрежной части Тауйской губы в трех районах: бух. Старая веселая и бух. Нагаева и мыса Нюкля. Пробы планктона отбирались сетью «Джеди» (площадь входного отверстия 0,1 м2, ячея фильтрующего конуса 0,168 мм) в слое 10-0 м на трех станциях в каждом районе с пятидневной дискретностью в одной и той же фазе прилива (по полной воде). Собранные пробы фиксировались смесью 70 % этилового спирта и формалина в пропорции 1:1. Камеральную обработку гидробиологических проб (всего 207) проводили по методике [10]. После идентификации личинок мидий в пробе [20] просчитывали их количество в камере Богорова, пересчитывали на 1 м3 водного объема и определяли среднедекадную численность в каждом районе. Все личинки двустворок измеряли тотально под бинокуляром с точностью до 25 мкм. Температуру регистрировали на глубине 2 м от поверхности воды. Статистическая обработка данных проводилась с помощью стандартных компьютерных программ. Сравнивались значения коэффициентов корреляции температуры воды и количества экземпляров личинок в планктоне. Данные по температуре воды и количеству личинок мидии достоверны при P< 0,05.
Результаты
Первые личинки мидий в планктоне прибрежной части Тауйской губы появились 26 июня при температуре поверхности воды 13,7-14,0ºС в количестве 5-8 экз/м3 (рис.1).
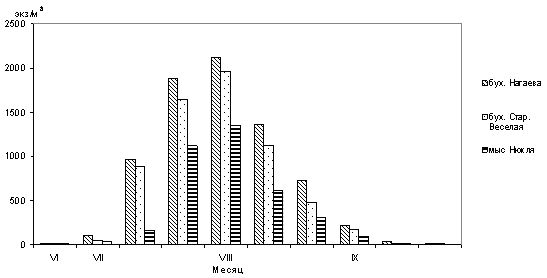
Рис.1. Сезонные изменения численности личинок M. trossulus (среднедекадные данные) в различных районах Тауйской губы в период с июня по сентябрь 2009 г.
К концу первой декады июля в планктоне находилось от 48 до 97 экз/м3 ранних велигеров c длиной раковины от 100 до 200 мкм. Во второй декаде июля численность личинок увеличилась до 963 экз/м3 и появились первые готовые к оседанию велигеры с длиной раковины 250-300 мкм, при этом продолжались многочисленные поступления в планктон ранних велигеров (рис. 2).
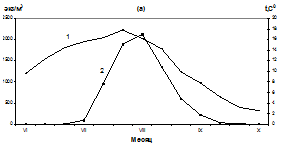
В третьей декаде июля - начале августа наблюдался резкий подъем численности личинок мидии. Максимальное количество – 2125 экз/м3 отмечалось в бух. Нагаева при средней температуре поверхности воды 16,2ºС (рис. 2 а). В районе бух. Старая Веселая максимальное количество личинок (1775 экз/м3) зарегистрировано в первой декаде августа при температуре воды 16,8ºС (рис. 2 б), в районе мыса Нюкля - 1330 экз/м3 при средней температуре поверхности воды 15,6ºС (рис. 2 б). Во второй декаде августа численность личинок мидий в планктоне снизилось, по-видимому, в результате охлаждения воды и оседания крупных велигеров. В третьей декаде августа их количество в планктоне колебалось от 673 экз/м3 - в районе бух. Старая Веселая и до 210 экз/ м3 в районе мыса Нюкля. В сентябре численность личинок в планктоне продолжало сокращаться до единичных особей к концу месяца. В октябре личинки мидии в планктоне не встречались.
Бух. Нагаева
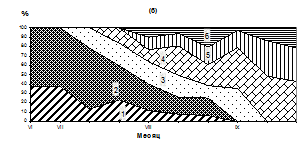
Бух. Старая Весёлая
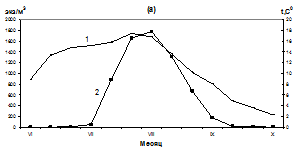
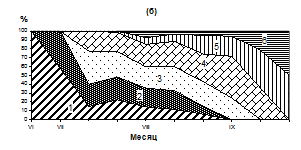
Мыс Нюкля
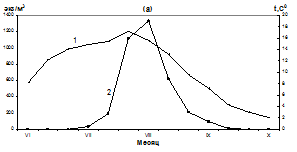
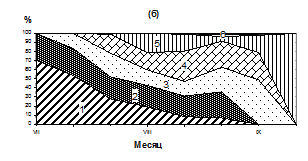
Рис. 2. Сезонные изменения температуры (1, а), численности (2, а) и размерного состава личинок M. trossilus (б) (среднедекадные данные). 1- 100-150 мкм; 2- 150-200 мкм; 3 – 200-250 мкм; 4 – 250-300 мкм; 5 - 300-350 мкм; 6 – 350-400 мкм.
В работах [9; 11; 3; 18] отмечалось, что тихоокеанская мидия приобретает способность оседать на субстрат при длине раковины не менее 250-270 мкм, а при длине 215 -230 мкм у личинки развивается пигментный глазок. По нашим данным в Тауйской губе глазок был в наличии у 75 % велигеров размером 230 мкм. Только при длине раковины 250 мкм глазок был практически у всех личинок мидий. Этот факт мы использовали при выделении доли личинок, теоретически готовых к оседанию в размерной группе более 250 мкм.
Таким образом, резкий подъем численности велигеров M. trossilus в конце июля – начале августа, зарегистрирован в период максимального прогрева воды в прибрежье Тауйской губы. 30 июля при температуре в поверхностном слое воды 17,5ºС отмечался массовый выход в планктон стерробластул мидий диаметром 75 мкм. Через пять дней стерробластулы перешли в стадию ранних велигеров с длиной раковины 100 мкм и более.
Массовый нерест мидий продолжался с 25 июля по 5 августа. В этот период несколько тысяч стерробластул постепенно переходили в раннюю стадию велигеров, которые впоследствии были обнаружены в планктоне при средней численности около 1000 экз/м3. Такое снижение численности, вероятно, связано с их элиминацией и разносом приливо-отливными течениями. В третьей декаде августа наблюдается охлаждение поверхностных вод до 9,6º С и численность личинок мидий в планктоне резко снижается (рис. 2, а, б). В этот период в планктоне преобладают велигеры размером 250-350 мкм, велигеры меньших размеров (150-200 мкм) составляют 10-25 % от общего количества личинок, а ранние велигеры (100-150 мкм) встречаются редко - от 1 до 6 % в зависимости от района.
В сентябре наблюдается очень быстрое выхолаживание наиболее мелководных прибрежных зон [17]. В планктоне преобладают крупные личинки мидий, численность, которых постоянно уменьшается вследствие оседания. В октябре личинки мидий уже не встречаются.
Обсуждение результатов
М. trossilus (Gould, 1850) – самый массовый моллюск на мелководьях северной части Охотского моря, обитает на поверхности грунта и занимает большие площади в прибрежной акватории Тауйской губы от литорали до сублиторали [16]. Массовые поселения – мидиевые банки располагаются от среднего до нижнего горизонта литорали. [8]. В 2009 г. массовый нерест мидий проходил в относительно сжатые сроки с 25 июля по 5 августа при прогреве воды свыше 17ºС. Однако в целом нерест был растянут, свидетельством чего было длительное присутствие велигеров М. trossilus в планктоне. Максимальная численность личинок мидии в горизонте 10-0 м составляла около 2125 экз/м3. В тоже время в прибрежной зоне, вблизи мидиевых банок в момент массового размножения максимальная численность личинок мидий достигала более 10000 экз/м3. При переходе личинок мидий из одной стадии в другую их численность снижалась на порядок. Наибольшие потери происходили при развитии D-велигеров из стереобластул (из 10 тысяч и более оставалось максимально 1000-1200 экз/м3 велигеров). По данным Милейковского (1961) из 1500 велигеров до стадии педивелигеров доживает максимум 200 экз/м3.
Таким образом, сроки появления и пребывания личинок в планктоне, их количество, степень растянутости нереста связаны со скоростью прогрева мест обитания родительских форм, последовательным вступлением в нерест особей, обитающих в пределах локальной акватории, и заносом личинок течениями из других районов. В целом, индивидуальная продолжительность личиночного развития у М. trossilus составляет в Тауйской губе примерно 35-40 суток, а личинки в планктоне в водах исследуемых районов регистрировались на протяжении 3 месяцев.
Следует добавить, что в бух. Нагаева и Старая Веселая в августе отмечалось большое количество крупных личинок мидий, находящихся в стадии оседания, причем их максимальные размеры были выше, чем в этот же период в районе мыса Нюкля. В литературе есть данные, указывающие на то, что личинки мидии могут задерживать метаморфоз при отсутствии подходящего субстрата для оседания [21; 15]. Так, в прибрежной части мыса Нюкля отмечалось присутствие илисто- песчаных грунтов и высока вероятность того, что основная масса личинок погибает, не найдя подходящего субстрата. В то же время в бух. Нагаева и Старая веселая гораздо больше подходящих для оседания субстратов. В этих бухтах ландшафт представлен в основном скальными грядами, валунами и галькой. В мелководной части скальные породы покрыты зарослями водорослей и морскими травами [7; 8].
В целом концентрации личинок тихоокеанской мидии в бух. Нагаева и Старая Веселая в течение всего исследуемого периода были более чем достаточными для успешного сбора спата. По нормативным данным, район следует считать перспективным для сбора спата мидии, если численность личинок в планктоне не менее 500 экз/м3 [5; 6]. Сравнительно невысокая концентрация личинок в районе мыса Нюкля обусловлена ее открытостью, активным воздействием на грунт ветрового волнения, небольшим количеством родительских форм и активным гидродинамическим режимом этого района.
Заключение
Результаты планктонных съемок показали, что в 2009 г. личинки тихоокеанской мидии присутствовали в планктоне в Тауйской губе с 25 июня до начала второй декады сентября. Динамика их численности характеризовалась наличием одного ярко выраженного максимума, регистрируемого в начале августа. Интервал между временем появления ранних личинок мидии в планктоне и моментом начала их оседания на субстрат составляет около 35-40 суток. Оседание личинок начиналось со второй декады августа, когда численность личинок в планктоне размером 250-350 мкм превышала 534 экз/м3. С 21 августа по 20 сентября количество личинок в планктоне готовых к оседанию резко сокращается от 374 экз/м3 до 7 экз/м.3 В третьей декаде сентября личинки мидии встречались только на двух станциях, по всей вероятности это связано с заносом их благодаря активным гидродинамическим режимом в этот период года. Полученные результаты позволяют утверждать, что послеличиночное расселение и оседание молоди тихоокеанской мидии на субстрат – является характерной особенностью биологии М. trossilus в Тауйской губе.
На основании полученных результатов, выявлено, что районы бух. Нагаева и Старая Веселая являются наиболее пригодными не только для сбора спата, но и для выставления подвесных коллекторов для выращивания тихоокеанской мидии.
Литература:
1. Агарова И. Я. Результаты многолетних наблюдений за популяцией Mytilus edulis L. на одной из литоральных отмелей Восточного Мурмана // Промысловые двустворчатые моллюски-мидии и их роль в экосистемах. Л. 1979а. - С. 8-10.
2. Беэр Т. Л. Личинки двустворчатых моллюсков в меропланктоне Великой Салмы Кандалакшского залива Белого моря. Океанология. – 2000. - Т.40. - № 5. - С. 714-719.
3. Брыков В. А., Семенихина О. Я., Колотухина Н. К. Динамика численности личинок мидии Mytilus trossilus в планктоне и их оседание на коллекторы в заливе Восток Японского моря // Биология моря. 2000. - №4. – С. 248-253.
4. Буяновский А. И., Куликова В. А. Распределение личинок мидии обыкновенной в планктоне и их оседание на коллекторы в заливе Восток Японского моря. // Биол. моря. 1986. - № 6. - С. 52-56.
5. Временная инструкция по биологии культивирования тихоокеаской устрицы. Владивосток: ТИНРО. 1984. – 25 с.
6. Временная инструкция по биологии культивирования съедобной мидии. – Владивосток: ТИНРО. 1988. – 36 с.
7. Галанин Д. А. Прибрежные сообщества беспозвоночных и водорослей макрофитов Берингова и Охотского морей (На примере Анадырского залива и Тауйской губы). Анадырь: НИЦ «Чукотка» ДВО РАН. 1997. - 24 с.
8. Жарников В. С. Размерно-возрастная и количественная характеристика мидии тихоокеанской Mytilus trossulus Gold, 1850 на литорали бух. Нагаева Тауйской губы. // Геология, география и биологическое разнообразие Северо-Востока России: Материалы Дальневосточной региональной конференции, посвященной памяти А.П. Васьковского и в честь его 95-летия. Магадан: СВНЦ ДВО РАН. 2006. - С. 343-347
9. Касьянов В. Л., Крючкова Г. А., Куликова В. А., Медведева Л.А. Личинки двустворчатых моллюсков и иглокожих. – М. : Наука, 1983. - 215 с.
10. Куликова В. А., Колотухина Н. К. Пелагические личинки двустворчатых моллюсков Японского моря. Методы, морфология, индентификация: Предприн. №. 21 Владивосток: ИБМ ДВО АН СССР. 1989. - 60 с.
11. Куликова В. А., Колотухина Н. К. Распределение пелагических личинок некоторых промысловых двустворчатых моллюсков в северо- восточной части залива Петра Великого // Экосистемные исследования: прибрежные сообщества залива Петра Великого. - Владивосток: ДВО АН СССР. 1991. – С. 99 -110.
12. Милейковский С. А. Личинки морских донных беспозвоночных и их роль в биологии моря. – М. : Наука, 1985. – 119 с.
13. Милейковский С. А. Некоторые проблемы экологии размножения морских донных беспозвоночных с пелагическим развитием // Тр. Мурман. морск. Биол. ин-та Ан СССР. - 1961. - Т.3 (7). - С. 147-169.
14. Полякова С. А. Динамика численности личинок приморского гребешка, тихоокеанской устрицы и тихоокеанской мидии в Амурском и Уссурийском заливах (Япоское море).// Изв. ТИНРО. - 2002. - Т.131. - С. 409-422.
15. Раков В. А. Биологические основы культивирования тихоокеанской устрицы Crassostrea gigas (Thunberg), в заливе Петра Великого: автореф. дис. … канд. биол. наук. – Владивосток, 1984. – 24 с.
16. Регель К. В. Морские и солоноватоводные беспозвоночные Тауской губы Охотского моря // Биологическое разнообразие Тауйской губы Охотского моря. - Владивисток: Дальнаука, 2005.- С. 479-521.
17. Чернявский В. И., Радченко Я. Г. Физико-географическая характеристика Тауйской губы Охотского моря. // Кн. Биологические основы развития лососеводства в Магаданском регионе. - Спб, 1994. – С.10-24.
18. Шепель Н. А. Биологические основы культивирования съедобной мидии в южном Приморье // Биол. моря. 1986. - № 4. - С. 14-21.
19. Шепель Н. А. Биология и культивирование мидии обыкновенной // Культивирование тихоокеанских беспозвоночных и водорослей. – М. : Агропромиздат. 1987. - С. 85-90.
20. DeBoyd L, Smith and Kevin B. Johnson. Мarine Coastal Plankton and Мarine Invertebrate Larvae. 1996. - P. 164.
21. Bayne B.L. The biology of mussel larvae // Мarine, mussels: their ecology and physiology. L: Cambridge Univ. Press. 1976. – P. 81-120.
