





Клостридии, эшерихии и стафилококки – нормальные обитатели кишечника человека, животных и птиц. Клостридии – облигатные анаэробы, а эшерихии и стафилококки — факультативные.
Клостридии, эшерихии и стафилококки являются полезными микроорганизмами, они синтезируют ряд витаминов — В, К, Е. Эшерихии и стафилококки являются антагонистами клостридий, тем самым уравнивают численность клостридий в организме, чтобы не развилась острая инфекция, в то же время наличие определенного числа клостридий обуславливает иммунизирующую субинфекцию, что необходимо для защиты организма от клостридий, попавших из внешней среды, при ранениях или других повреждениях, тем самым предупреждая развитие инфекции.
Среди множества эшерихий и стафилококков — антагонистов клостридий – окажется несколько десятков клеток E. coli, стафилококков, которые лишены свойства антагонизма, именно они совокупляются с клостридиями.
У эшерихий энергия размножения превышает другие бактерии, обитателей кишечника, что дает им возможность за короткий срок изменить численность в свою пользу, вызывая геморрагическое воспаление слизистой оболочки кишечника, результатом чего является кровавая диарея.
Эшерихии, имеющие свойства токсигенности, являются возбудителями многих других заболеваний — перитонита, менингита, токсиноинфекции и др.
Объекты внешней среды: почва, вода, продукты питания, овощи загрязняются выделениями животных, в которых во множестве содержатся клостридии, эшерихии, стафилококки. Особенно загрязняется почва; если создаются условия, способствующие их сохранению — рН 7,2–7,6, наличие питательных веществ, температура; указанные микроорганизмы начинают развитие и даже размножение.
Попадание этих бактерий в организм человека и животных в большинстве случаев оральное, для клостридиев также характерно попадание через открытые раны; т. к. с частицами земли попадают клостридии, которые находят в ране подходящие условия для развития и размножения — в первую очередь анаэробные условия, во вторую — наличие гликогена, который является обязательным для питания и размножения клостридий.
Спаривание клостридий и эшерихий возможно и в объектах внешней среды, если температура, влажность, рН, питательный субстрат находятся на оптимальном уровне.
Эшерихии никогда не будут патогенными, если не произойдет их скрещивание с клостридиями. При этом клостридии передают эшерихиям токсигенные, гемолитические, антигенные, сахаролитические свойства, устойчивость к антибиотикам; эти же свойства клостридии передают и стафилококкам. Этим мы разрешили вопрос патогенности эшерихий и стафилококков. Все началось с того, что ученые пытались культивировать анаэробные клостридии в аэробных условиях, для того, чтобы было легче изготавливать вакцину.
Известно, что клостридии не синтезируют фермент каталазу, который необходим для разрушения перекиси водорода, накапливающейся в питательной среде, являющегося ядром для анаэробов, которые не продуцируют каталазу, а аэробы и факультативные анаэробы синтезируют данный фермент. Мы задались целью путем скрещивания передать данное свойство клостридиям. Эшерихии не смогли выполнять роль донора, однако они оказались довольно хорошими реципиентами, а клостридии — донорами.
Таким образом была раскрыта суть патогенности эшерихий, клостридий: Cl. oedematiens, Cl. perfringens, Cl. septicum, Cl. chauvoei передавали эшерихиям и стафилококкам токсичность, антигенность, гемолитическую активность, сахаролитические свойства, устойчивость к антибиотикам.
Передачу токсигенных свойств контролировали заражением подопытных животных — кроликов, морских свинок, белых мышей; нейтрализацию токсина — реакцией нейтрализации; антигенные связи устанавливали реакцией агглютинации, с использованием гипериммунной сыворотки; гемолитическую активность — засевом глюкозо-кровяного агара; резистентность к антибиотикам — внесением в питательную среду антибиотиков.
Передачу токсигенных свойств стафилококкам от клостридий устанавливали заражением подопытных животных в брюшную полость (белые мыши, морские свинки) и кроликам введением внутрикожно культур рекомбинантов, также реакцией плазмокоагуляции, гемолитическую активность — засевом в глюкозо-кровяной агар.
В любом случае, как с эшерихиями, так и со стафилококками имела место передача плазмидной ДНК от клостридий. Кроме этих тестов засняли взаимно связанные клетки — клостридий с эшерихиями, клостридий со стафилококками в электронном микроскопе, где очень хорошо видна связь донора (клостридии) с реципиентами (эшерихии, стафилококки), иногда слияние бактериальных оболочек; в любом случае переносится плазмидная ДНК.
Вели также скрещивание клостридий со стафилококками при низкой температуре. При этом нарушается клеточная оболочка, и плазмидная ДНК легко покидает клетку донора и переносится в клетку реципиента с помощью конъюгационного мостика или слиянием клеточных оболочек.
Ниже представлены полученные электронно-микроскопические снимки:
- Конъюгация между клетками Cl. perfringens и E. coli. Клетка большего размера Cl. perfringens (донор), меньшего размера — E. coli (реципиент); они связаны между собой конъюгационным мостиком. Этим мостиком переносится плазмидная ДНК Cl. perfringens в клетку E. coli. Соответственно E. coli приобретает патогенные свойства, гемолитическую активность, антигенность, резистентность к антибиотикам и др.
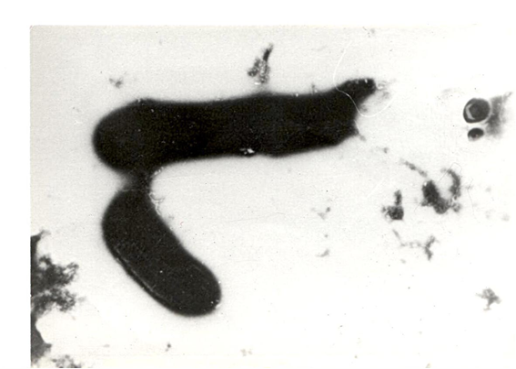
- Конъюгация между клетками Cl. septicum и E. coli; клетка меньшего размера (справа) — донор Cl. septicum; они связаны между собой конъюгационным мостиком, с которого переносится в клетку реципиента плазмидная ДНК, соответственно в ней детерминированные признаки — токсигенность, гемолитические свойства и др.
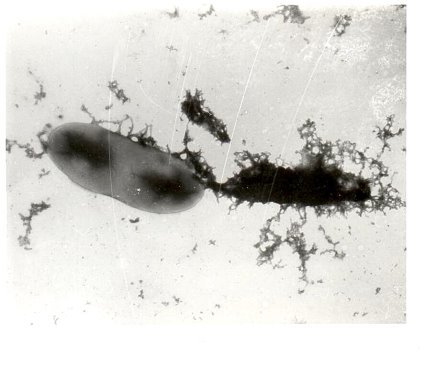
- Конъюгация между Cl. oedematiens и E. coli. Клетки меньшего размера, сферической формы — E. coli (реципиент). Между донором (Cl. oedematiens) и реципиентом (E. coli) связывающий мостик, с которого в одном направлении переносится (с донора на реципиент) плазмидная ДНК, вместе с ней — указанные свойства.

- Те же микробы Cl. oedematiens и E. coli. Сравнительно меньшего размера — E. coli; они связаны между собой конъюгационным мостиком; также имеет место слияние оболочек. В любом случае осуществляется перенос плазмидной ДНК

- Конъюгация между Cl. chauvoei и E. coli. Клетка меньшего размера — E. coli. Хорошо виден соединяющий мостик — происходит перенос ДНК от донора (Cl. chauvoei) к реципиенту (E. coli)
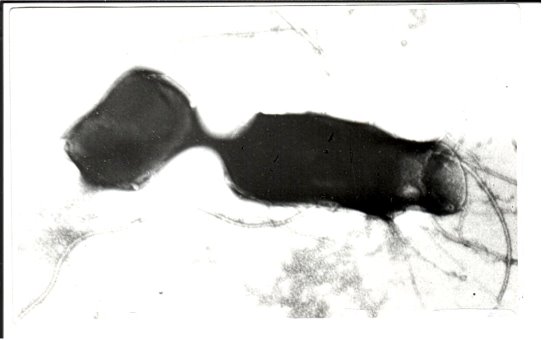
- Конъюгация между Cl. chauvoei и E. coli. Клетка меньшего размера — E. coli. (реципиент). Между ними несколько соединительных мостиков, которые переносят разные плазмиды, тем самым различные наследственные признаки (см. выше).

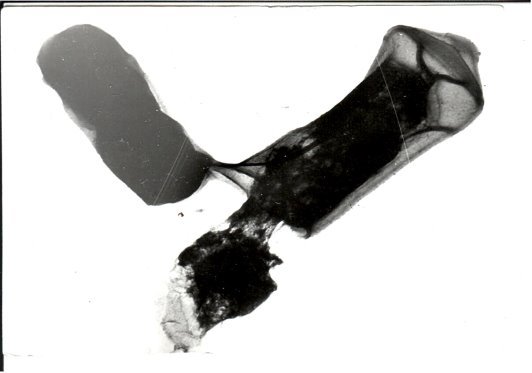
- Конъюгация между клетками Cl. perfringens. Связь между ними устанавливается слиянием клеточной оболочки. Мы зафиксировали перенос множественной резистентности к антибиотикам (R -фактор). Донор, несущий R –фактор, по-видимому, клетка, находящаяся сверху. Размножение начинается после того, как между клетками устанавливается связь (слияние клеточных оболочек). По литературным данным известно, что прокариоты размножаются поперечным делением, по всей вероятности, это неверно.
У животных организмов для сохранения потомства и наследственных признаков обязательным условием является установление связи между мужскими и женскими клетками, без этого невозможно размножение. В популяции клеток первоначально осуществляется дача импульса от донора к реципиенту (ДНК донора дает импульс ДНК реципиенту), и только после этого начинается поперечное деление.
Всевышний именно так определил продолжение рода — универсальный закон всего живого.
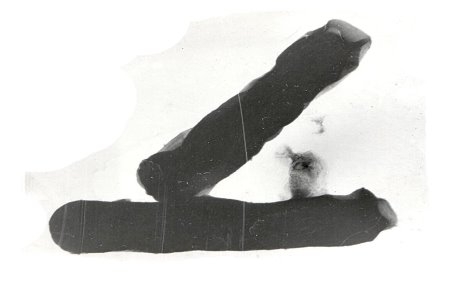
- Конъюгация между Cl. perfringens и стафилококками — донор (Cl. perfringens), реципиент (Staphylocjccus). Их связывает конъюгационный мостик. Вокруг донора несколько клеток реципиента, что указывает на наличие в клетке Cl. perfringens нескольких плазмидных ДНК, которые распределяются в клетках реципиента; переносятся в клетки стафилококков плазмиды, несущие синтез экзотоксинов, гемолитической активности, антигенность, резистентность к антибиотикам, сахаролитические свойства и др.
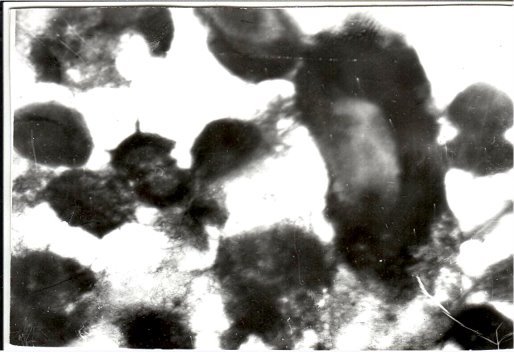
- Как на 8-м снимке Cl. perfringens (донор) связана со стафилококками (реципиент) таким образом, что одна клетка донора обладает притягательной силой для нескольких реципиентов клеток, которым передает плазмидную ДНК, несущую те признаки, которые указаны выше, с некоторыми конъюгационным мостиком, с другими — слиянием оболочек.
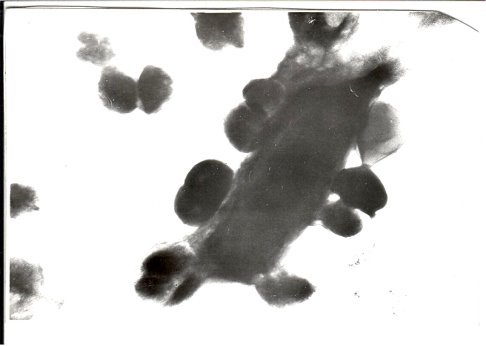
- Клетки сферопластов и протопластов (Cl. perfringens — донор и стафилококки — реципиент) спариваются при низкой температуре. Предварительно мы получали из Cl. perfringens и стафилококков сферопласты и протопласты, затем инкубировали смешанно при низких температурах. Передавалась гемолитическая активность и резистентность к антибиотикам; плазмида, несущая токсигенность, элиминирована.
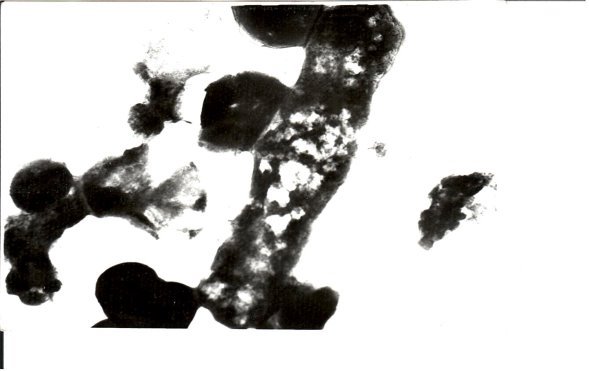
- Неполные протопласты (Cl. perfringens — донор и стафилококки реципиент) спаривались при низкой температуре; передается плазмиднаяДНК и вместе с ней гемолитическая активность и резистентность к антибиотикам.
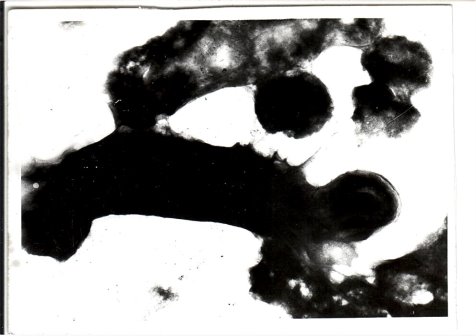
- Донор (Cl. perfringens) полностью лишен клеточной стенки — находится в состоянии протопласта. Стафилококки (несколько клеток) частично сохранили клеточную оболочку (сферопласты). Их спаривание происходит при низкой температуре, при этом переносится с донора на реципиент гемолитическая активность и резистентность к антибиотикам.
Хочу обратить Ваше внимание на следующее — протопласты не изменили свою форму, по литературным данным бактерии в экстремальных условиях изменяют форму, — переходят в оргонтные формы. Например, шаровидные — неверно. Конъюгация между ними идет.
Доноры соединяются с реципиентом конъюгационным мостиком или сливаются безоболочные клетки — донора и реципиента.
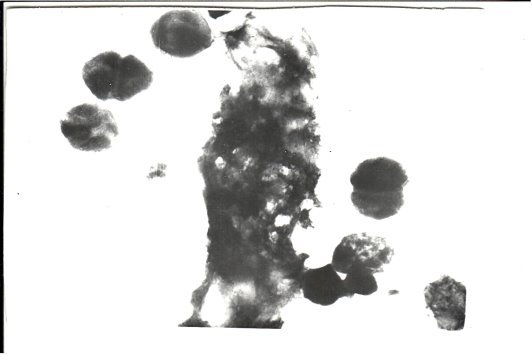
- В организме человека и животных на бактериальную клетку действуют ферменты, а также низкая температура внешней среды, которые разрушают оболочку. Низкая температура обуславливает разрушение клеточной оболочки, в данном случае у Cl. perfringens. Это в свою очередь вызывает выход цитоплазмы наружу, т. е. в клетку макроорганизма (не всегда), тем самым освобожденная хромосома включается в хромосому макроорганизма, в результате происходит синтез белка с измененными свойствами, что, иногда, вызывает опухолевый рост.
Индукцию опухолевого роста мы получили на белых мышах протопластами и лизитами токсигенных клостридий, также рекомбинантами эшерихий и стафилококков, что представляет, в одном случае, свободную молекулу ДНК, которая находится в лизате, в другом случае — высвобожденную из протопластов и клеток рекомбинантов ДНК.
Таким образом, опухолевый рост находится в причинной связи с токсигенными клостридиями, рекомбинантами эшерихий и стафилококков, которым передается от клостридиев плазмидная ДНК (Дж. Начкебия, Е. Начкебия, К. Начкебия — Известия аграрной науки, 2007, т.5, № 1, с.129–131. Тбилиси).
- R-форма колоний Cl. perfringens на глюкозо-кровяном агаре — плоские, шероховатые, морщинистые с зазубренными краями, хорошо выраженным гемолизом. Эти колонии — стрептомицинорезистентные мутанты, лишенные вирулентности, получены путем многократного пассирования на средах с антибиотиком; впоследствии использовались в опытах по трансформации.
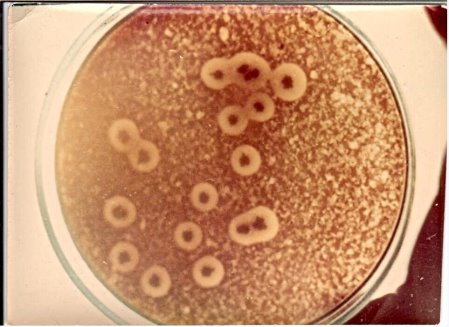
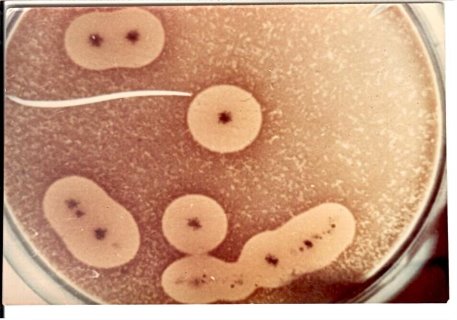
- R-форма колоний Cl. perfringens на сахарном агаре; здесь колонии плоские, шероховатые, морщинистые с зазубренными краями. Они лишены вирулентности; использовались в опытах по трансформации.
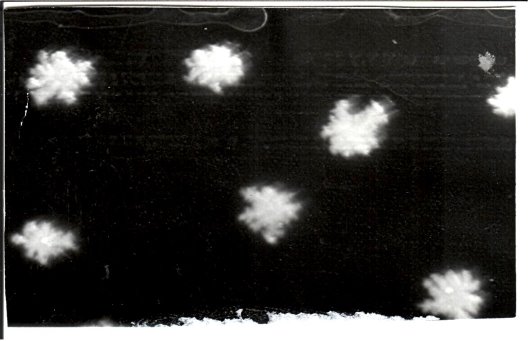
- ДНК, выделенная из стрептомицинорезистентного варианта Cl. perfringens. ДНК осаждена этанолом (верхний слой), переносится стеклянной палочкой в жидкость следующего состава: 0,15 М NaCl и 0,015 М цитрат Na; ДНК моментально растворяется.

- Исходная культура Cl. perfringens — клетка без воздействия стрептомицина (ультраструктура)
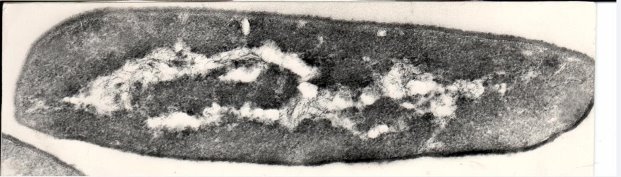
- Ультраструктура трансформированного варианта Cl. perfringens. Перенос плазмидной ДНК стрептомицинорезистентности с трансформированного варианта на исходную культуру.
Таким образом, доказана возможность получения гибридных штаммов с полезными свойствами, в результате скрещивания токсигенных анаэробных микроорганизмов с авирулентными эшерихиями, сообщающая при этом патогенные, гемолитические и антигенные свойства, характерные для клостридий. Попутно разработана методика селекционирования рекомбинатов E. coli из организма белых мышей, инфицированных смесью клостридий с эшерихиями; также разработана методика скрещивания анаэробных микроорганизмов с эшерихиями и стафилококками в физиологическом растворе NaCl.
Из рекомбинантных штаммов E. coli изготовлена вакцина и получена гипериммунная сыворотка против клостридиозов и эшерихиозов.
Из эталонных штаммов токсигенных клостридий изготовлена вакцина и получена гипериммунная сыворотка против колибактериоза и анаэробных инфекций.
Полученные нами результаты исследований по вопросам влияния токсигенных анаэробов на патогенные и другие свойства эшерихий и стафилококков позволяют глубже раскрыть механизм появления новых серогрупп E. coli и стафилококков с токсигенными и гемолитическими свойствами, могут способствовать более основательному изучению внутриклеточных процессов, протекающих после включения в них новых детерминант, направленных на получение штаммов с полезными свойствами; способствуют раскрытию механизма взаимодействия разных видов микроорганизмов в ассоциации в естественных средах их обитания и на этой основе появлению патогенных вариантов.
На основе проведенных исследований раскрывается механизм возникновения тяжелых кишечных заболеваний, токсико-септических инфекций, колиэнтеритов, пищевых отравлений и многое другое.
Установление факта передачи токсигенности эшерихиям и стафилококкам от патогенных клостридий внесет коренные изменения в вопросы диагностики, лечения и профилактики колиинфекций и стафилококковых заболеваний; внесет ясность во внехромосомные наследственности эшерихий, стафилококков и анаэробных микроорганизмов, тем самым открывая широкие возможности генетического моделирования при решении теоретических проблем молекулярной биологии.
Результаты исследований коренным образом изменяют имеющиеся до сих пор представления о патогенности эшерихий и стафилококков и дают совершенно новое направление исследованиям в области инфекционной патологии и генетики бактерий.
Литература:
- Начкебия Д. В., Начкебия Е. Д. Получение протопластов клостридий, эшерихий и стафилококков при низких температурах //Межгос. научный сборник по актуальным вопросам ветеринарии. Баку-Гянджа, 1996.
- Начкебия Д. В., Начкебия Е. Д. Слияние протопластов клостридий со стафилококками при низких температурах //Межгос. научный сборник по актуальным вопросам ветеринарии. Баку-Гянджа, 1996.
- Начкебия Д. В., Начкебия Е. Д., Шаматава Т. В. Приготовление суспензии конъюгирующих бактерий для электронной микроскопии // Межгос. научный сборник трудов.Ч.II. Тбилиси, 1997, с. 73–75.
- Начкебия Д. В., Начкебия Е. Д. Совместное замораживание клостридий с эшерихиями и стафилококками // Межгос. научный сборник трудов.Ч.II. Тбилиси, 1997, с. 75–78.
- Начкебия Д. В., Начкебия Е. Д., Начкебия К. Д. Причинная обусловленность патогенности эшерихий, связанная с совместным обитанием их токсигенными клостридиями (сообщение 1) //Известия аграрной науки, т.4. Тбилиси, 2004, с.93–96.
- Начкебия Д. В., Начкебия Е. Д., Начкебия К. Д. Причинная обусловленность патогенности эшерихий, связанная с совместным обитанием их токсигенными клостридиями (сообщение 2) //Известия аграрной науки, т.3.№ 2. Тбилиси, 2005, с.143–146.
- Начкебия Д. В., Начкебия Е. Д., Начкебия К. Д. Причинная обусловленность патогенности эшерихий, связанная с совместным обитанием их с токсигенными клостридиями (сообщение 3) //Известия аграрной науки, т.3, № 3. Тбилиси, 2004, с.195–197.
- Начкебия Д. В., Начкебия Е. Д., Начкебия К. Д. Причинная обусловленность патогенности эшерихий, связанная с совместным обитанием их с токсигенными клостридиями (сообщение 4) //Известия аграрной науки, т.3, № 4. Тбилиси, 2004, с.118–123.
- Начкебия Д. В., Начкебия Е. Д., Начкебия К. Д. Токсигенные клостридии как причинный фактор патогенности эшерихий //Известия аграрной науки, т.4, № 2. Тбилиси, 2006, с.122–126.
- Начкебия Д. В., Начкебия Е. Д., Начкебия К. Д. Опухолевый рост, индуцированный белым мышам протопластами и лизатами токсигенных клострдий и клостридиальными рекомбинантами эшерихий и стафилококков //Известия аграрной науки, т.5, № 1. Тбилиси, 2007, с.129–131.
- Начкебия Д. В., Начкебия К. Д., Начкебия Е. Д. Влияние токсигенных клостридий на биологические свойства эшерихий и стафилококков //Известия аграрной науки, т.9. Тбилиси, 2011, с.97–104.
- Начкебия Д. В., Начкебия К. Д., Начкебия Е. Д. Метод выделения клостридиальных рекомбинантов из сердца павших белых мышей //Известия аграрной науки, т.10, № 4. Тбилиси, 2012, с.100–103.
- Начкебия Д.В Влияние токсигенных анаэробов на биологические свойства эшерихий и стафилококков //Автореферат диссертации доктора ветеринарных наук, 1992. 53 с.
