





В статье автор рассматривает особенности регуляции биосинтеза белка у прокариот и эукариот, а также факторы, влияющие на него.
Ключевые слова: биосинтез белка, прокариоты, эукариоты, факторы, система оперон, белки, ферменты, белок-репрессор.
В нашем мире существует 2 вида организмов: прокариотические и эукариотические. Они отличаются строением и происходящими в них биологическими процессами, в том числе и биосинтезом белка — разновидностью пластического обмена, в ходе которого генетическая информация ДНК преобразуется в последовательность аминокислот будущего белка. На этот сложный процесс у про- и эукариот оказывают влияние разные факторы.
Регуляция биосинтеза у прокариот происходит на уровне транскрипции, поскольку он неразрывен с трансляцией. Он основан на регуляции активности генов, схема которой была открыта в 1961 Жакобом и Моно и названа опероном. Ученые проводили опыты на примере кишечной палочки и описали действие лактозного оперона. Они определили, что гены функционально неодинаковы: различают структурные гены, которые составляют структуру оперона, кодируют последовательность аминокислот в белке, и регуляторные гены, влияющие на работу структурных генов посредством белка-репрессора, который он кодирует. Любой оперон строится по определенному принципу: вначале стоит промотор, ответственный за связывание с РНК-полимеразой, за ним — оператор, включающий и выключающий структурные гены и определяющий работу гена-регулятора, далее следует последовательность генов, кодирующих определенную структуру белка, и заканчивается оперон геном-терминатором, сигнализирующем о прекращении синтеза РНК [4].
Рассмотреть действие оперона проще всего на лактозном опероне кишечной палочки (Рис. 1). При наличии глюкозы в цитоплазме бактерии, белок-репрессор находится на промоторе, тем самым препятствуя началу транскрипции. При попадании путем пассивного транспорта в клетку лактозы, последняя связывается с белком-репрессором, что приводит к отсоединению белка от промотора. На данном участке становится возможным транскрипция, в результате образуется и-РНК, кодирующая 3 белка. После трансляции и-РНК мы получаем фермент β-галактозидазу, пермиазу и β-галактозид трансацетилазу. Первый расщепляет лактозу на глюкозу и галактозу, второй является интегральным мембранным белком, обеспечивающим легкое и быстрое проникновение лактозы из окружающей среды в клетку. Когда лактоза заканчивается, она отщепляется от белка-рецептора и тот возвращается на промотор, препятствуя синтезу и-РНК. Таким образом, на примере работы оперона наиболее ярко видна регуляция биосинтеза белка в прокариотической клетке.
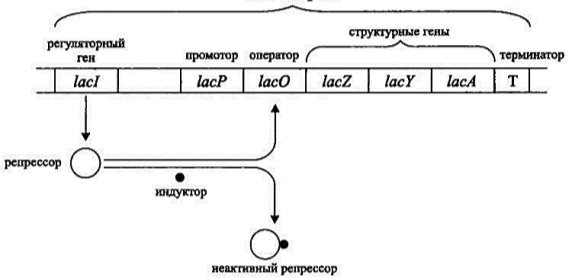
Рис. 1. Лактозный оперон
Другой пример — триптофанный оперон кишечной палочки. Он работает по другому принципу. До тех пор, пока триптофан расходуется, клетка его синтезирует. Но как только возникает избыток данной аминокислоты, она связывается с белком-репрессором. Данный белок присоединяется к промотору оперона, что приводит к прекращению синтеза триптофана. Со временем клетка расходует запасенную аминокислоту, начинает испытывать недостаток. Тогда триптофан отсоединяется от белка-репрессора, последний больше не может удерживаться на промоторе и освобождает. После этого начинается синтез и-РНК с матрицы оперона, после трансляции которой клетка получает необходимую ей кислоту триптофан. Цикл повторяется [2].
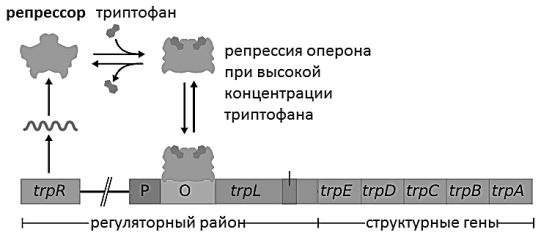
Рис. 2. Триптофанный оперон
Вообще, биосинтез микроорганизмов может осуществляться двумя путями: быстрым и более медленным. В первом случае речь идет о секундах или минутах, а механизм регуляции активности фермента зависит от изменений молекул, входящих в его состав. Во втором случае процесс может занимать часы, а количество фермента определяется интенсивностью процессов синтеза и распада оного. В обоих случаях работает принцип обратной связи — благодаря низкомолекулярным соединениям определяется количественное соотношение различных веществ в составе клетки, а структура и количество этого соединения определяется регуляторными механизмами биосинтеза [3].
В эукариотических клетках процессы транскрипции и трансляции разделены ядерной мембраной: транскрипция происходит в ядре, трансляция — в цитоплазме, поэтому в качестве одного из этапов биосинтеза выделяют экспорт из ядра. В пределах эукариотических клеток также можно рассматривать систему оперон, однако она будет иметь свои особенности.
Первая особенность состоит в пространственной организации оперона: если у эукариот гены оперона находятся последовательно друг за другом, то в эукариотической клетке репрессор может находится не в том же участке хромосомы, что промотор и структурные гены, а приближаться к ним только в интерфазу при высоком уровне компактизации хромосомы [1].
Другая особенность — не только репрессоры играют роль в регуляции синтеза белка. Существуют участки, влияющие на скорость транскрипции генов. Энхансеры являются усилителями транскрипции, поскольку способны связывать регуляторные белки, активирующие этот процесс. Их антагонисты — сайленсеры — так называемые «глушители»; они связываются с определенными белками, чем замедляют процесс. Считается, что энхансеры и сайленсеры способствуют изменению топологии цепей ДНК, чем приближают промотор к последовательности генов, которую необходимо транскрибировать.
Такой способ регуляции считается достаточно экономичным, однако на него затрачивается большое количество времени, поэтому регуляция белка у клеток данного типа осуществляется на уровне трансляции посредством регуляторных веществ, присоединяющихся к 3’ и 5’ не транскрибируемым участкам. Здесь количественное соотношение белков регулируется благодаря разной продолжительности жизни м-РНК, определяемой полиадениловым фрагментом 3’ конца ДНК. Кроме того, структура и-РНК определяет сродство к рибосомальным субъединицам, что влияет на длину полисомы, а значит и на количество образующегося белка. Возможно также полное подавление трансляции при действии неблагоприятных и стрессовых факторов, которые негативно сказываются на факторах инициации.
Таким образом, у прокариот и эукариот механизмы регуляции биосинтеза белка различны. Для первого типа организмов характерна регуляция на уровне транскрипции, для второго — на уровне и транскрипции, и трансляции. В обоих случаях применима схема оперона, однако существуют определенные различия для этих видов клеток: важно учитывать трехмерную организацию оперона и наличие энхансеров и сайленсеров в эукариотической клетке. Количественное содержание белков в этом типе клеток находится под влиянием регуляторных веществ и определяется генами ДНК.
Литература:
- Галицкий В. А. Возникновение эукариотических клеток и происхождение апоптоза // Цитология. 2005. Т. 47. Вып. 2. С. 103–120.
- Коничев А. С., Севастьянова Г. А. Молекулярная биология. — Издательский центр «Академия», 2012. — 400 с.
- О.-Я. Л. Бекиш. Медицинская биология. — Витебск: Ураджай, 2000. — С. 53.
- Сергеев В. Н., Нолл Э. X., Заварзин Г. А. Первые три миллиарда лет жизни: от прокариот к эвкариотам // Природа. 1996. № 6. С. 54–68.
