





Пусть неизвестный генотип по одному признаку 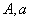 скрещивается с доминантной особью, и его потомок опять скрещивается с доминантной и так далее. Надо найти распределение вероятностей каждого из возможных генотипов в
скрещивается с доминантной особью, и его потомок опять скрещивается с доминантной и так далее. Надо найти распределение вероятностей каждого из возможных генотипов в 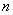 -ом поколении (см.
-ом поколении (см. ). Как известно, составлен таблица вероятностей перехода к возможным генотипам (различные строки — это вероятности возможных генотипов скрещиваемого, имеющим генотип
). Как известно, составлен таблица вероятностей перехода к возможным генотипам (различные строки — это вероятности возможных генотипов скрещиваемого, имеющим генотип 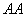 , столбцы — возможные вероятности генотипов потомства):
, столбцы — возможные вероятности генотипов потомства):
|
|
|
|
|
|
|
1 |
0 |
0 |
|
|
|
|
0 |
|
|
0 |
1 |
0 |
Таблица означает, что генотип при скрещивании с генотипом 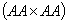 даёт потомок с вероятностью 1, с
даёт потомок с вероятностью 1, с 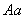
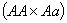 даёт потомок с вероятностью
даёт потомок с вероятностью 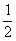 , с
, с 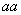
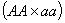 даёт потомок с вероятностью о и так далее (первый столбец).
даёт потомок с вероятностью о и так далее (первый столбец).
Обозначим через 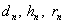 , соответственно, вероятности генотипа (доминантный), (гитрозиготный), (рецессивный),
, соответственно, вероятности генотипа (доминантный), (гитрозиготный), (рецессивный), 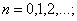 и
и 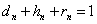 .
.
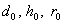 являются исходными вероятностями неизвестной особи, соответственно.
являются исходными вероятностями неизвестной особи, соответственно.
Для получение вероятностей 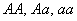 в потомстве суммируем вероятности этого генотипа при различных возможных генотипах неизвестной особи (по столбцу таблицы). Для получение вероятности генотипа в первом поколении берём вероятность получения этого генотипа при условии, что генотип неизвестной особи
в потомстве суммируем вероятности этого генотипа при различных возможных генотипах неизвестной особи (по столбцу таблицы). Для получение вероятности генотипа в первом поколении берём вероятность получения этого генотипа при условии, что генотип неизвестной особи 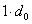 , и прибавим вероятностной особи , т. е.
, и прибавим вероятностной особи , т. е. 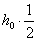 . Тогда вероятность получения в первом поколении при скрещивании равно
. Тогда вероятность получения в первом поколении при скрещивании равно 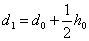 .
.
Аналогично 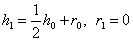 .
.
Для получения вероятностей следующего поколения мы воспользуемся с той же таблицы.
Получим: 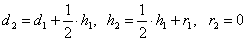 .
.
Поставляя значения 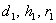 , имеем
, имеем
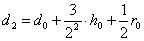 ,
, 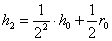 ,
, 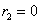 .
.
Аналогично, для –го поколения скрещивания с , имеем:
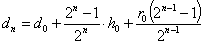 ,
, 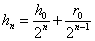 ,
, 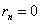 .
.
Отсюда следует теорема:
ТЕОРЕМА-1. 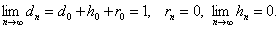
Если неизвестный особь скрещивается с особью с генотипом и в результате вероятность того, что получается равно: 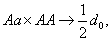
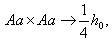
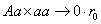 , суммируя имеем:
, суммируя имеем:
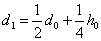 .
.
Аналогично, вероятность того, что имеем генотип , равно
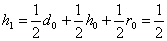 ,
,
а для генотипа
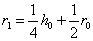 .
.
Для второго поколения
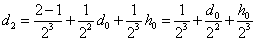 ,
,
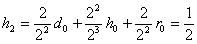 ,
,
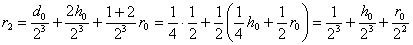 .
.
Для -ного поколения скрещивания с имеем
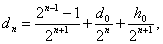
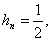
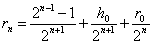 .
.
Отсюда следует
ТЕОРЕМА-2. 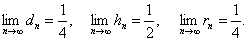
Если неизвестный особь скрещивается с особью с генотипом и в результате вероятность того, что в -ом поколении получается, соответственно, генотипы , , равно
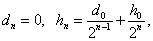
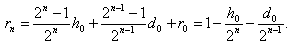
Тогда имеет место:
ТЕОРЕМА-3. 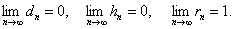
В частности, при 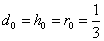 приведем таблицу для
приведем таблицу для 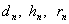 .
.
|
n |
1 |
2 |
3 |
||||||
|
d(n) |
h(n) |
r(n) |
d(n) |
h(n) |
r(n) |
d(n) |
h(n) |
r(n) |
|
|
1 |
0,5 |
0,5 |
0 |
0,25 |
0,50 |
0,25 |
0 |
0,5 |
0,5 |
|
2 |
0,75 |
0,25 |
0 |
0,25 |
0,50 |
0,25 |
0 |
0,25 |
0,75 |
|
3 |
0,875 |
0,125 |
0 |
0,25 |
0,50 |
0,25 |
0 |
0,125 |
0,875 |
|
4 |
0,9375 |
0,0625 |
0 |
0,25 |
0,50 |
0,25 |
0 |
0,0625 |
0,9375 |
|
5 |
0,96875 |
0,03125 |
0 |
0,25 |
0,50 |
0,25 |
0 |
0,03125 |
0,96875 |
|
6 |
0,984375 |
0,015625 |
0 |
0,25 |
0,50 |
0,25 |
0 |
0,015625 |
0,984375 |
|
7 |
0,9921875 |
0,0078125 |
0 |
0,25 |
0,50 |
0,25 |
0 |
0,0078125 |
0,9921875 |
|
8 |
0,99609375 |
0,00390625 |
0 |
0,25 |
0,50 |
0,25 |
0 |
0,00390625 |
0,99609375 |
|
9 |
0,998046875 |
0,00195313 |
0 |
0,25 |
0,50 |
0,25 |
0 |
0,001953125 |
0,998046875 |
|
10 |
0,999023438 |
0,00097656 |
0 |
0,25 |
0,50 |
0,25 |
0 |
0,000976563 |
0,999023438 |
Заключение: Из таблицы видно, что неизвестный особь при скрещивание с имеет вероятность 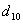 , очень близкое единице, следовательно,
, очень близкое единице, следовательно, 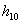 близко к нулю, если скрещивание происходит с , то и
близко к нулю, если скрещивание происходит с , то и 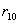 близки к
близки к 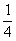 , при скрещивании с близко к нулю,
, при скрещивании с близко к нулю, 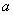 близко к единице.
близко к единице.
Литература:
1. Нейман Ю. Вводный курс теории вероятностей и математической статистики. Из. «Наука», М. 1968.
2. Математика и естествознание, Из. «Просв»., М. 1969.
3. Marek Kimmel, David E. Axelrod. Branching Processes in Biology. Springer. 2002.
